
坦鲷及其饲养

独一无二的 Cyphotilapia frontosa(六间)
来源:摘自《CICHLIDS NEWS》(2019年第1期)
(翻译:ChatGPT-4o)
Cyphotilapia frontosa概述
Cyphotilapia frontosa是Lake Tanganyika较深岩石栖息地中常见的物种。这种雄伟的鱼类可长至超过35 cm(14英寸)的总长度,几乎在任何有岩石的栖息地都能找到它们,通常出现在20米(66英尺)或更深的水域。幼鱼会在稍浅一些的水层活动。实际的深度因种群和栖息地而异:在Kavala群中的Kibige岛,可以在5至40米(16-130英尺)深度范围内发现大量的C. frontosa,而在Kapampa,该物种能被观测到的最浅深度约在25至30米(80-100英尺)之间。C. frontosa以群体生活,常见到一条大型雄鱼“负责”整群,带领数条雌鱼(有时正在口孵)和一两条体型较小的雄鱼一起活动的情形并不少见。
C. frontosa动作相当迟缓,似乎从不慌张。幼年个体主要以软体甲壳类动物为食,但成体多半是肉食鱼类,主要捕食其他鱼类。它们在咽骨上的牙齿又尖又细,与那些主要捕食大型无脊椎动物、拥有臼齿状牙齿的慈鲷形成鲜明对比。对野生个体胃容物的研究发现,其主要食物残骸来自鱼类以及贻贝(Poll 1956),还包括昆虫幼体、虾和蜗牛(Büscher 2011)。在人工饲养环境中的观察也进一步证明了C. frontosa属于食鱼性:它确实能够——而且确实会——吞食与自身体长几乎相当于其一半大小的缸友。
也许C. frontosa最引人注目的特征就是其头顶的隆起。雄鱼最为明显,但雌鱼并非完全没有这种突起。这道隆起由背部肌肉(非脂肪)构成,向前延伸,可能在配对识别或威慑其他雄鱼时发挥作用。体型较大的雄鱼通常隆起更明显,不过Mbita 岛种群的雄鱼似乎在体型还较小时就会出现明显的隆起。
C. frontosa以一种十分独特的方式繁殖,这在除Trematochromis benthicola之外的其他母系口孵慈鲷中都未见过。繁殖前,雄鱼的蓝色体色会明显加深,尤其在吻部。雄鱼不会挖掘产卵场或防守领地。当一雄一雌准备产卵时,它们并不会离开群体,而是共同寻找一处合适地点,随后雄鱼会对该地点进行弱势防御。雄鱼不会像其他慈鲷那样颤动或摆动身体,而是鳍折拢,以缓慢移动的方式在选好的场地“示范”给雌鱼看。雄鱼完成这一“示范巡游”后,雌鱼也会以相似的方式进入场地并产下一枚卵。但雌鱼并不会像其他慈鲷那样转身去捡起刚产下的卵,而是向后退去将其收入口中。完成拾卵后,它并未让出位置给雄鱼,而是继续产下更多卵。如果不受干扰,雌鱼可以连续“摆动”5到6次,一共产下多达20枚卵,然后才会离开产卵场,让雄鱼再次进入。极有可能雄鱼释放的精液具有相当的“持久力”,即使在排出数分钟后才与卵接触,仍能让这些卵受精。人们从未观察到雌鱼会像其他口孵慈鲷那样用嘴轻触雄鱼的臀鳍以摄入精液。当雌鱼离开场地后,雄鱼会再次带领雌鱼回来,并重复相同过程,直到所有卵都产完。雄鱼似乎从不追赶雌鱼。卵在口中被孵育约5周后,小鱼才被释放出来。

对物种的错误认知
C. frontosa在整个湖区都有分布,并出现了几个明显不同的地理变异体。1906年,Boulenger基于在Kigoma采集的模式标本首次描述了该物种。Kigoma种群的主要特征是身体上有6条横纹、头部1条横纹,颊部呈深蓝色,且背鳍中带有可变的黄色。在玩家中,这一形态也被称为“7-bar Frontosa”。
在2003年,Takahashi & Nakaya描述了该属中的第二个物种,命名为Cyphotilapia gibberosa。其模式标本采集于湖泊最南端的Kasenga Point(Zambia的Mpulungu附近)。该种群的特征是在躯干部分拥有5条横纹、头部1条横纹。一般会以为躯干5条和6条横纹之间的区别足以简单地区分两个物种,但奇怪的是,Takahashi & Nakaya并未采用这一显而易见的区别,而是声称这两者的区别在于上下侧线之间的鳞片数:C. frontosa有2行,而C. gibberosa有3行。他们还将他们的新物种与分布在最北端的布隆迪种群进行比较,并宣称即使布隆迪形态也有5条躯干横纹与1条头部横纹(与C. gibberosa相同),但它与Kigoma形态(7-bar Frontosa)同种。所以,条纹的区别被排除,而以鳞片的行数为主。
Genevelle(2004)并不认同这种观点,认为布隆迪种群代表了第三个物种,并暂时将其称作Cyphotilapia sp. '6-bar frontosa'。到了2007年,Takahashi(即最初C. gibberosa描述的主要作者)与Ngatunga及Snoeks一同重新检视了来自北部的6-bar Cyphotilapia的分类地位。他们检查了来自8个不同地点的32个标本,与C. frontosa的正模标本以及21个来自模式产地Kigoma的新鲜采集标本进行比较。基于此次大范围研究,他们得出了这样的结论:“……在形态测量与计数性状之间确实存在显著差异;然而,因为这些性状在这些形态(C. frontosa & 6-bar)之间存在大量重叠,所以我们认为它们属于同种。”于是,我们又回到了两个物种:C. frontosa和C. gibberosa——而6-bar frontosa被归入C. frontosa(也就是包含7-bar的那个)。Kigoma种群身体上多出的那条纹被再次视为无关紧要,这一次得到了更多鱼类学家的认同。

Tawil(2010)指出Takahashi等(2007)出现了错误(并未解释错误之处何在),并认为出现“中间型”的原因并非地理变异,而是杂交。然而,在缺少DNA研究证明杂交的情况下,必须有明确存在的亲本物种才行,而事实并非如此,否则我们早就能确凿无疑地证明这是两个不同的物种。实际上,在这个问题上DNA分析确有可能提供决定性证据:只要给所谓的亲本物种都定义一个DNA标记,然后检验中间种群是否同时拥有两者标记(如果是杂交起源),或并无二者混合(如果不是杂交)。过去确实采集过DNA研究所需的样本,但迄今并没有任何研究发表过这方面的结论,更没有证据明确表明我们正在面对的物种数量是多于1种的。
有意思的是,Takahashi等(2007)也对来自北部的标本与26个C. gibberosa标本进行比较,仍以上下侧线之间的鳞片数——他们曾用来区分C. frontosa与C. gibberosa的首要特征——为依据。而他们的表格显示,一些C. gibberosa标本只有2行鳞片(理应是3行),而有些C. frontosa标本反倒是3行(本应是2行)。换句话说,Takahashi & Nakaya(2003)当初规定的区分两个物种的主要特征也存在重叠,因此并不能作为准确的鉴定依据。很显然,C. gibberosa只是C. frontosa的一个地理变异,虽然在各地演化出了若干地方性变异,但还没有哪一种变异真正演变到不能被其他变种识别、从而形成一个新物种的程度。

Genevelle(2004)的一些发现反而可以支持此观点:据报道,Malagarasi河三角洲以南(即7-bar C. frontosa分布区最南端以南)所采集到的Cyphotilapia标本拥有3行侧线间鳞片,理论上应被归为C. gibberosa,但其体色模式更像布隆迪形态的C. frontosa。Genevelle并没有由此得出“这是处在C. frontosa和推定C. gibberosa之间的中间形态”这一合乎逻辑的结论,反而提出这是该属的第四个物种:C. sp. 'Centre-Est'。尤其对不熟悉野外慈鲷多样性的水族爱好者而言,每当看到进口商鱼缸里出现一个新“形态”,往往会倾向于将其视为一个新物种。
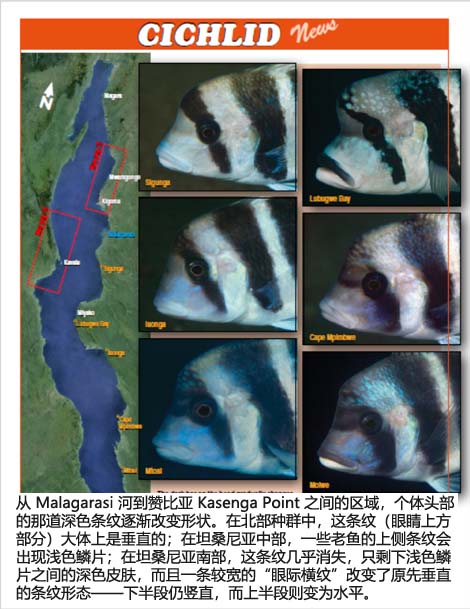
我曾经在Malagarasi河与坦桑尼亚-赞比亚边境之间的50多个地点潜水,并在大部分地点拍摄了Cyphotilapia。从地图中可以看到,从北往南,这些Cyphotilapia的头部花纹是一个从明显的竖直条带逐渐演变至带有网状黑边的浅色鳞片,最终在更往南的地方演变成一条清晰的眶间带的连续变化过程。
尽管Takahashi & Nakaya(2003)声称C. gibberosa只分布于湖泊南半部(西岸在Moba以南,东岸在Miyako以南),但在Kalemie以北的Kavala 岛也有3行侧线间鳞片的Cyphotilapia,因此理论上也应被归为C. gibberosa,或者如果按照“分裂主义者”的思路,干脆再设立一个第五个物种:C. sp. 'Centre-Ouest'!我在Kibige和Bilila岛上发现了“3行鳞片”形态,而在Kabimba及更北部地区的种群则是2行侧线间鳞片,因此它们被归为C. frontosa。

通过研究慈鲷 DNA 中的所谓 SINEs(短散在重复元件),Kasuhiko Takahashi(并非为 C. gibberosa 命名的那位 Takahashi)及其同事(1998)发现,Cyphotilapia 这一支系(即坦噶尼喀湖慈鲷家族谱系中的一个分支)大约在 1,000 万年前就已出现,并且自那时起便独立于目前湖中所有其他慈鲷支系。这并不意味着物种 C. frontosa 本身就有 1,000 万年历史——虽然也不排除这种可能性——而是说与之相同支系的祖先在 1,000 万年前就已存在。Cyphotilapia 在分化出来之前,与 lamprologine 有共同祖先,但自那以后 lamprologine 已经演化成 8 个属,现有约 85 个物种;同期出现的 Ectodini 与 Haplochromini 分别演化出了约 40 个和 1500 个物种,而 Cyphotilapia 这一支系却仅保留在一个分支上。
在数个研究团队对 Cyphotilapia 进行分子(DNA)和形态学检验之后,Salzburger 等(2002)将这种坦噶尼喀湖高头慈鲷划入一个单型的新族,命名为 Cyphotilapiini。随后 Muschick 等(2012)和 Meyer 等(2015)又将 Trematochromis benthicola 视作该族的第二个物种。这些结果再次表明,虽然其他支系(族)在漫长岁月里演化出了众多属和物种,Cyphotilapia 似乎并没有经历类似的大分化。
在这几乎无法想象的漫长时期里(1,000 万年——当时既没有黑猩猩,也没有大猩猩,更没有人类),湖泊水位多次发生波动,许多时候水位下降到将湖泊一分为三的程度。对马拉维湖的水位研究显示,在过去的 130 万年间,下降 200 米(约 656 英尺)或更多的情形就出现了至少 24 次(Lyons 等,2015)!人们经常将坦噶尼喀湖的此类湖面大幅度波动与新物种的产生联系起来,认为在三大“古湖”中被隔离的种群在湖面上升后与近缘“表亲”重新汇合时,依旧保持各自独立。(然而,在一些从未因湖面下降而被分隔的湖泊中——比如马拉维湖和维多利亚湖——慈鲷显然也有能力在不到百万年的时间里演化出数千种物种。)但 Cyphotilapia 在这一时期似乎并未分化为多个物种,因为今天我们在湖中任何地点都只发现同一个物种——C. frontosa。

出于提升销量的目的,鱼类出口商常给每批鱼都起不同名字,企图让鱼友们以为这是新物种。有一位赞比亚的出口商曾在同一捕捞点采集所有的 C. frontosa,将它们运回后在养殖池里先行适应,再将“蓝色” frontosa 与“普通” frontosa分开,因为蓝色个体能卖更高价。实际上,“蓝色”只是在同一群体里随机出现的体色差异,特别是濒临繁殖的个体,常比群里其他鱼展现出更多蓝色。当然,蓝色 frontosa 的流行也与 1990 年代初从刚果(当时称扎伊尔)南部水域出口的 frontosa 有关。这些鱼来自水深较大的区域,有的深达 60 米(约 197 英尺),似乎深水环境下的个体体色更偏蓝。你如果查看那张展示各地理变异的分布图,会发现它们的纹路多少有所差异,因此明智的做法是只购买确切采集点来源的鱼,以保持品系纯正。(编辑注:不过,出口商的信息常常并不可靠!)祝各位好好享受你们的慈鲷!
参考文献
--Boulenger,G.A. (1906)。《对坦噶尼喀湖鱼类学的第四次贡献--W.A. Cunnington 博士在 1904-1905 年第三次坦噶尼喀考察期间收集鱼类的报告》。伦敦动物学会论文集,17(6): 537-619。
--Büscher,H.H. (2011)。《观察野生Cyphotilapia的摄食行为》,德国慈鲷协会信息,42(7): 150-154。
--Genevelle,E. (2004). 《Cyphotilapia属的分布、生态和收集》Regan, 1920。慈鲷科, 4: 53-66.
--Lyons, R.P., C.A. Scholz, A.S. Cohen, J.W. King, E.T. Brown, S.J. Ivory, T.C. Johnson,A.L. Deinof, P.N. Reinthal, M.M. McGlue, & M.W. Blome. (2015). 《东非130万年连续水文气候记录及其对进化和生物多样性模式的影响》。美国国家科学院院刊,112(51):15568-15573。
--Meyer, B.S., M. Matschiner & W. Salzburger (2015). 《基于基因组多标记方法的坦噶尼喀湖慈鲷部落级系统发育》。分子系统学与进化,83: 56-71。
--Muschick,M.,A. Indermaur,W. Salzburger(2012年)。《慈鲷适应性辐射内的趋同进化》。当代生物学,22(24): 2362-2368。
--Poll,M. (1956). 鲈形目:慈鲷科。 《坦噶尼喀湖水生物勘探(1946-1947 年)》-科学成果, 3(5B): 1-619。
--Salzburger,W.、A. Meyer、S. Baric、E. Verheyen 和 C. Sturmbauer(2002 年)。《坦噶尼喀湖慈鲷种群的系统发育及其与中非和东非Haplochromine慈鲷鱼群的关系》。 系统生物学,51(1): 113-135。
--Takahashi, K., Y. Terai, M. Nishida, & N. Okada (1998)。《来自慈鲷的一个新的短穿插重复元件(SINEs)家族:SINEs 在同源基因座上的插入模式支持坦噶尼喀湖慈鲷四大类群的单系化观点》。分子生物学与进化,15(4): 391-407。
--Takahashi, T. & K. Nakaya (2003)。《来自非洲坦噶尼喀湖的Cyphotilapia新种(鲈形目:慈鲷科)》。Copeia、2003(4): 824-834。
--Takahashi, T., B. Ngatunga, & J. Snoeks (2007)。《来自非洲坦噶尼喀湖的Cyphotilapia frontosa(鲈形目:慈鲷科)六条形态的分类地位》。 鱼类学研究,54(1): 55-60。
--Tawil, P. (2010)。《Cyphotilapia:坦噶尼喀湖的加冕慈鲷》。慈鲷资讯, 19(3): 20-28。

--沪ICP备16038729号-1-- | Powered By DedeCMS