C. frontosa以一种十分独特的方式繁殖,这在除Trematochromis benthicola之外的其他母系口孵慈鲷中都未见过。繁殖前,雄鱼的蓝色体色会明显加深,尤其在吻部。雄鱼不会挖掘产卵场或防守领地。当一雄一雌准备产卵时,它们并不会离开群体,而是共同寻找一处合适地点,随后雄鱼会对该地点进行弱势防御。雄鱼不会像其他慈鲷那样颤动或摆动身体,而是鳍折拢,以缓慢移动的方式在选好的场地“示范”给雌鱼看。雄鱼完成这一“示范巡游”后,雌鱼也会以相似的方式进入场地并产下一枚卵。但雌鱼并不会像其他慈鲷那样转身去捡起刚产下的卵,而是向后退去将其收入口中。完成拾卵后,它并未让出位置给雄鱼,而是继续产下更多卵。如果不受干扰,雌鱼可以连续“摆动”5到6次,一共产下多达20枚卵,然后才会离开产卵场,让雄鱼再次进入。极有可能雄鱼释放的精液具有相当的“持久力”,即使在排出数分钟后才与卵接触,仍能让这些卵受精。人们从未观察到雌鱼会像其他口孵慈鲷那样用嘴轻触雄鱼的臀鳍以摄入精液。当雌鱼离开场地后,雄鱼会再次带领雌鱼回来,并重复相同过程,直到所有卵都产完。雄鱼似乎从不追赶雌鱼。卵在口中被孵育约5周后,小鱼才被释放出来。
对物种的错误认知
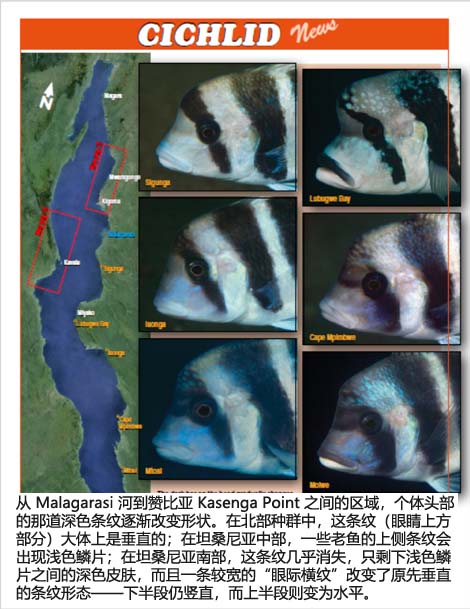
C. frontosa在整个湖区都有分布,并出现了几个明显不同的地理变异体。1906年,Boulenger基于在Kigoma采集的模式标本首次描述了该物种。Kigoma种群的主要特征是身体上有6条横纹、头部1条横纹,颊部呈深蓝色,且背鳍中带有可变的黄色。在玩家中,这一形态也被称为“7-bar Frontosa”。